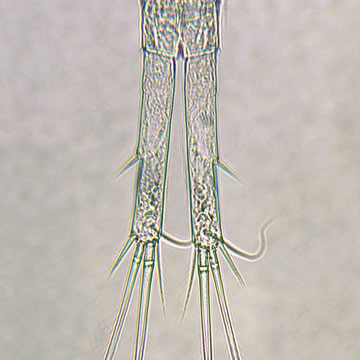
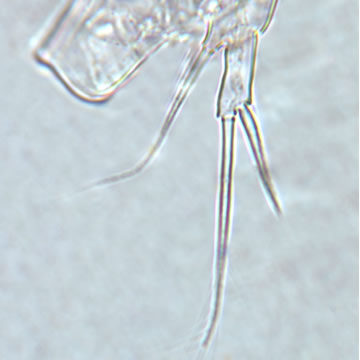
| |
|||||||||||||||||||
|
|||||||||||||||||||
| Ecology and Behavior | |||||||||||||||||||
| Characteristics | |||||||||||||||||||
|
|||||||||||||||||||
| Similar Species | |||||||||||||||||||
| D. thomasi has been previously referred to as Diacyclops bicuspidatus thomasi (1). In 1882, S.A. Forbes first described this species as Cyclops thomasi; his son, E.B. Forbes, later listed the species as Cyclops bicuspidatus. In 1933, Gurney combined the two as a distinct North American subspecies, Cyclops bicuspidatus thomasi. Later Yeatman (1959), included this taxon in the subgenus Diacyclops. Reed (1963), Dussart (1969), and Kiefer (1978) determined that lake-dwelling Diacyclops bicuspidatus thomasi is distinct from the temporary pond species D. bicuspidatus and then conferred full-species status as Diacyclops thomasi (1). | |||||||||||||||||||
| Geographic Distribution & Reported Habitats | |||||||||||||||||||
| D. thomasi is widely distributed throughout North America and is the most common copepod in zooplankton samples in the Great Lakes (1). Examples of specific reported locations include Lake Michigan (2), Belews Lake, North Carolina (3), Oneida Lake, New York (4), Lake Ontario (5), and Quimet and Conneli Lakes, Montreal, Canada (6). A Great Lakes (USA) study determined D. thomasi was found at least 0.3 m off the bottom 75% of the time; this indicates most individuals are planktonic rather than epibenthic (residents at or near the bottom) (1). |
|||||||||||||||||||
| Food & feeding behavior | |||||||||||||||||||
| D. thomasi are herbivorous until the fourth instar (CIV) stage, after which they become omnivorous (4). Prey items are usually microplankton (15-100 µm), generally motile and lacking a test or protective covering (4)(8). In a laboratory study of prey selection, D. thomasi consistently selected the soft-bodied rotifers Synchaeta pectinata, Polyarthra major and P. remata (2). Other soft-bodied rotifers, the small P. vulgaris and Ascomorpha ecaudis, and the large adult Asplanchna priodonta were not preferred. Bosmina longirostris, Chydorus sphaericus and the loricate species Keratella cochlearis and K. crassa avoid predation by D. thomasi (2). The mucus sheath of Ascomorpha ecaudis serves as a defense against predation by D. thomasi (2). When D. thomasi encounters the sheath of the rotifer, the cyclopoid stops swimming and rapidly moves its feeding appendages attempting to remove the mucus protecting the rotifer (2). Late copepodid stages of D. thomasi prey on ciliates, rotifers, small cladocerans, juvenile copepods (4) and larval fish (7). D. thomasi partially consumes larger prey items and share large prey with other D. thomasi (1). |
|||||||||||||||||||
| Reproductive Habits | |||||||||||||||||||
| In the Great Lakes (USA), females begin producing clutches of 10-40 eggs in early spring with 2-3 generations per year (9)(10). Cultured D. thomasi eggs can reach adulthood within 28-35 days and females can produce a second clutch within four days after the first hatching (9). Eggs sampled from the Missouri River measured 68-79 µm diameter (1). |
|||||||||||||||||||
| Predation | |||||||||||||||||||
In the Great Lakes, D. thomasi is consumed by fish such as alewife, bass, bloaters, ciscoes, carpsuckers, perch, sculpin, shiners, whitefish and walleyes (11) (12). D. thomasi in Lake Ontario exhibited a significant population decline after the introduction of the invasive cladoceran, Cercopagis pengoi (5). |
|||||||||||||||||||
| Competition | |||||||||||||||||||
Immature omnivorous stages of D. thomasi compete with Daphnia for rotifer and ciliate prey (4). |
|||||||||||||||||||
| Migration | |||||||||||||||||||
In the North American Great Lakes adult D. thomasi exhibit diel vertical migrations (deep water during the day; migrating to surface at night) with the highest concentrations during daylight near or above the thermocline (13)(14)(15). |
|||||||||||||||||||
| Seasonal Variations | |||||||||||||||||||
In the Great Lakes (USA) between summer and early autumn 10-20% of D. thomasi were encysted in the sediments (16). During the clearwater phase (reduced phytoplankton biomass from grazing) in Lake Oneida, NY, D. thomasi disappeared from the water column but encysted as fourth instar copepodids (CIV) in the sediments(4)(6). The Lake Oneida population of copepodids emerged from encystment during the fall, primarily from the deep water (4). In contrast, diapausing D. thomasi (CIV stage), in Marion Lake, British Columbia, emerged from the sediments in spring (4). Diapause in D. thomasi is most likely triggered by a combination of photoperiod, temperature, and food limitation conditions typical of late spring in temperate zones (4). |
|||||||||||||||||||
| References | |||||||||||||||||||
|
(1) HUDSON, P. L., L. T. LESKO, J. W. REID, AND M. A. CHRISCINSK E. 2003. Cyclopoid copepods of the Laurentian Great Lakes. Ann Arbor, MI. Great Lake Science Center Home Page. http://www.glsc.usgs.gov/greatlakescopepods/Key.asp?GROUP=Cyclopoid (2) STEMBERGER, R. 1985. Prey Selection by the copepod Diacyclops thomasi. Oecologia. 65: 492-497. (3) MARCOGLIESE, D. J. AND G. W. ESCH. 1992. Alterations of Vertical Distribution and Migration of Zooplankton in Relation to Temperature. American Midland Naturalist. 128: 139-155. (4) HANSEN, A. M. AND N. G. HAIRSTON, JR. 1998. Food limitation in a wild cyclopoid copepod population: direct and indirect life history responses. Oecologia. 115: 320-330. (5) LAXSON, C. L., K. N. MCPHEDRAN, J. C. MAKAREWICZ, I. V. TELESH, AND H. J. MACISAAC. 2003. Effects of the non-indigenous cladoceran Cercopagis pengoi on the lower food web of Lake Ontario. Freshwater Biology. 48: 2094-2106. (6) BAUD, A., C. CUOC, AND V. ALEKSEEV. 2004. Ultrastructure of the digestive tract of Diacylops thomasi (Cyclopoida, Copepoda) during different stages of encystment during a summer diapause. Acta Zoologica. 85: 181-189 . (7) LEBLANC, J. S., W. D. TAYLOR, AND O. E. JOHANNSSON. 1997. The feeding ecology of the cyclopoid copepod Diacyclops thomasi in Lake Ontario. J. Great Lakes Res. 23: 369-381. (8) HARTIG, J. H., D. J. JUDE, AND M. S. EVANS. 1982. Cyclopoid predation on Lake Michigan fish larvae. Can. J. Fish. Aquat. Sci. 39: 1563-1568. (9) EWERS, L. A. 1930. The larval development of freshwater copepods. The Ohio State Univ. Frantz Theodore Stone Lab. Contr. No. 3: 1-43. (10) SELGEBY, J. H. 1975. Life histories and abundance of crustacean zooplankton in the outlet of Lake Superior, 1971-1972. J. Fish. Res. Board Can. 32: 461-470. (11) BALCER, M. D., N. L. KORDA, AND S. I. DODSON. 1984. Zooplankton of the Great Lakes: A guide to the identification and ecology of the common crustacean species. The University of Wisconsin Press, Madison. 174pp. (12) EWERS, L. A. 1933. Summary report of Crustacea used as food by the fishes of the western end of Lake Erie. Trans. Am. Fish. Soc. 63: 379-390. (13) WELLS, L. 1960. Seasonal abundance and vertical movements of planktonic crustacea in Lake Michigan. U.S. Fish and Wildlf. Serv. Fish. Bull. 60: 343-369. (14) CARTER, J. C. H. 1969. Life cycles of Limnocalanus macrurus and Senecella calanoides, and seasonal abundance and vertical distribution of various planktonic copepods in Parry Sound, Georgian Bay. J. Fish. Res. Board Can. 26: 2543-2560. (15) WILSON, J. B., AND J. C. ROFF. 1973. Seasonal vertical distributions and diurnal migration patterns of Lake Ontario crustacean zooplankton. Proc. 16th Conf. Great Lakes Res. 190-203. (16) NALEPA, T. F. 1985. Occurrence of a resting stage in cyclopoid and harpacticoid copepods in nearshore Lake Michigan. J. Great Lakes Res. 11: 59-66. |
|||||||||||||||||||
| Additional Pictures | |||||||||||||||||||
|
|||||||||||||||||||
| Quicktime Movies | |||||||||||||||||||
|
|||||||||||||||||||
| Barcode: Ribosomal DNA-28S D3 expansion segment | |||||||||||||||||||
|
|||||||||||||||||||