| |
|||||||||||||||||||
|
|||||||||||||||||||
| Ecology and Behavior | |||||||||||||||||||
| Characteristics | |||||||||||||||||||
|
|||||||||||||||||||
| Similar Species | |||||||||||||||||||
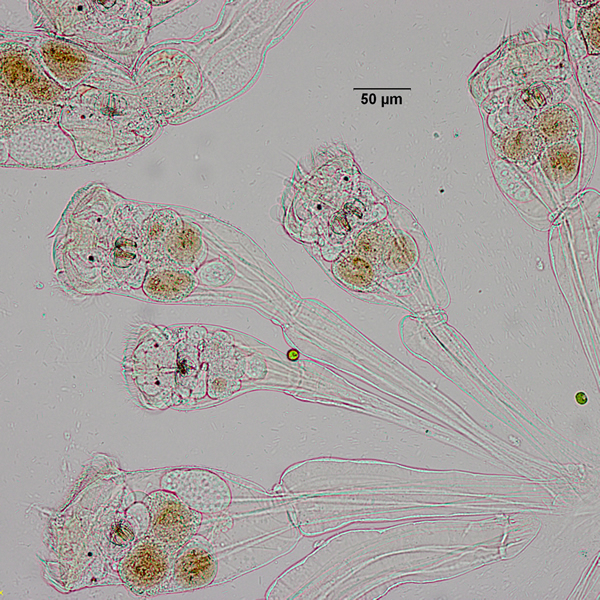
| C. hippocrepis is often found with C. unicornis, which has a similar colonial behavior; both forming Type II colonies (explained below) (1). C. unicornis is often more abundant and can withstand a wider variety of environmental conditions than C. hippocrepis. | |||||||||||||||||||
| Morphological Features | |||||||||||||||||||
| Type I colonies have a free-swimming immature stage that actively selects a substrate to settle on whereas type II colonies forgo the free-swimming stage and become directly incorporated into the parental colony (1). When C. hippocrepis colonies become too large, daughter colonies may split from the parental colony, or larvae may form a new colony (1). Because Type II colonies are continuously adding offspring, they can go on indefinitely throughout the season, while Type I colonies will come to an end if no new recruits settle (1). | |||||||||||||||||||
| Geographic Distribution | |||||||||||||||||||
| C. hippocrepis have been reported in a wide variety of lakes including, Lake Washington (4)(5), inland lakes of northern lower Michigan (2), Pleasant Lake, New Hampshire (1), South Lake, Main Lake, Northeast Arm, North Lake, Missisquoi Bay of Lake Champlain (7), a fishless pond of the Southern Andes (6), Laguna Fantasma (a small temporary pond near Nahuel Huapi lake, West Argentina) (6), and Estonian Lakes, particularly Lake Peipus (8)(9). | |||||||||||||||||||
| Reported Habitats | |||||||||||||||||||
| C. hippocrepis usually inhabit small fishless ponds or temporary pools (6), oligotrophic and mesotrophic lakes (8) and eutrophic lakes (9)(10). | |||||||||||||||||||
| Feeding Behavior and Competition | |||||||||||||||||||
| C. hippocrepis are grazers (10). Very small particles, possibly plastids and organelles from soft-walled algae, have been found in the stomachs of Conochilus (4). In addition, the diatom Melosira distans (5 µm x 13 µm), an empty chlorophyte cell wall (13 µm x 8 µm), and small green algae (5 µm in diameter) have also been reported within the stomachs of Conochilus (4).
Animals observed living together had a much higher survival rate than solitary animals. Combined w ater currents may enable greater food intake with less energy spent (11). Rate of food intake can be increased in this manner, allowing extra energy to be spent on repair and reproduction (1). This would contribute to the success of the colonies and be a positive selective force in their evolution. Edmondson (1943) wondered at what size a population would grow too large and survival would be negatively affected (11). Other researchers have hypothesized that colony formation is energy saving when applied as a predatory defense, but the increased colony size could reduce the filtering efficiency of each individual and prevent energy intake (1)(6)(11). If colony size increases too much, the collective filtering currents could interfere with the individual filtering, causing less food to be cleared and more energy spent (1). Although currents represent a trade-off in large colony formation, the gelatinous sheath encasing these rotifers may be more cost-effective than forming spines or other morphological defenses. Symbiotic algae, Phormidium and an unidentified chlorophyte, may grow as epibionts on the gelatinous matrix of Conochilus colonies (4). These epibionts may provide some nutritional benefits to Conochilus, as vegetative cells of these symbiotic algae have been found in the stomach contents of C. hippocrepis (4). |
|||||||||||||||||||
| Reproductive Habits | |||||||||||||||||||
| In temporary pool habitats, C. hippocrepis develop from resting eggs once the rains refill the pools (6). Eggs are rarely seen in the colony because development is so advanced when the eggs are laid that they hatch in less than an hour (1)(4). A mictic or sexual reproduction period, has not been detected; C. hippocrepis could be acyclic (sexual reproduction occurring irregularly) in some lakes (10). | |||||||||||||||||||
| Predation on Conochilus | |||||||||||||||||||
| Forming protective colonies is a unique strategy used by the entire genus Conochilus; this is an uncommon practice with other planktonic rotifers (6). Parabroteas sarsi, a large predacious calanoid copepod, is one of C. hippocrepis main predators. C. hippocrepis increases its colony size as P. sarsi also increases in size (6). As a result, the mean colony diameter is always larger than the maxilliped size of P. sarsi, thereby preventing predation of the entire colony (6). The gelatinous matrix provides protection from attack as C. hippocrepis can retract into the safety of the matrix becoming too large to be engulfed (3)(6). Asymmetrical colonies have been observed, and could be a result of either predation or a recent colony split (4). Leptodora populations increase with decreasing C. hippocrepis density; and when Leptodora is scarce, C. hippocrepis and C. unicornis became abundant (4). The assumption that Leptodora preys on C. hippocrepis and C. unicornis has no direct evidence because the transparency of Leptodora made observations of feeding difficult (4). C. hippocrepis trophi (mouth parts), 48 total, were reported within the stomach contents of a single Leptodora (4). The calanoid copepod, Epischura nevadensis is also a predator of C. hippocrepis, but their populations are not always affected by the presence of E. nevadensis (4). Other reported predators of Conochilus are Polyphemus pediculus, Cyclops scutifer, Mesocyclops edax and Diaptomus spatulocrenatus (12). | |||||||||||||||||||
| Competition | |||||||||||||||||||
| In Laguna Fantasma (41°S, 72°W, Argentina), the rotifer colonies reached maximum size and then disappeared from this temporary pond, at which time the Daphnia middendoriffiana population began to increase (6). Whether this large Daphnia caused C. hippocrepis to disappear is not known (6). | |||||||||||||||||||
| Additional Pictures | |||||||||||||||||||
 |
 |
 |
|||||||||||||||||
| References | |||||||||||||||||||
|
(1) WALLACE, R. L. 1987. Coloniality in the phylum Rotifera, Hydrobiologia. 147: 141-155. (2) STEMBERGER, R. S. 1990. An inventory of rotifer species diversity of northern Michigan inland lakes, Arch. Hydrobiol. 118: 283-302. (3) THORP, J. H. AND A. P. COVICH. 2001. Ecology and Classification of North American Freshwater Invertebrates, second edition, Academic Press. pp.1056. (4) EDMONDSON, W. T., AND A. H. LITT. 1987. Conochilus in Lake Washington. Hydrobiologia. 147: 157-163. (5) EDMONDSON, W. T. AND A. H. LITT. 1982. Daphnia in Lake Washington. Limnol. Oceanogr. 27: 61-62. (6) DIEGUEZ, M. AND E. BALSEIRO. 1998. Colony size in Conochilus hippocrepis: Defensive adaptation to predator size. Hydrobiologia. 387-388: 421-425. (7) Plattsburg State University of New York. 2004. A Guide to the Zooplankton of Lake Champlain, Lake Champlain Research Institute. Scientia Discipularum. 1: 38-66. (8) MAEMETS, A. 1983. Rotifers as indicators of lake types in Estonia. Hydrobiologia. 104: 357-361. (9) VIRRO, T., AND J. HABERMAN. 1993. The rotifers of Lake Peipus. Hydrobiologia. 255/256: 389-396. (10) VIRRO, T. 2001. Life cycle patterns of rotifers in Lake Peipsi. Hydrobiologia. 446/447: 85-93. (11) EDMONDSON, W. T. 1945. Ecological Studies of Sessile Rotatoria, Part II: Dynamics of Populations and Social Structures. Ecol. Monograph. 15: 141-172. (12) WILLIAMSON, C. E. 1983. Invertebrate predation on planktonic rotifers. Hydrobiologia. 104: 385-396. |
|||||||||||||||||||
| Quicktime Movies | |||||||||||||||||||