|
Home / Chlorophyceae / Unicells / Flagellated / Chlamydomonas |
||||
|
|
||||
|
|
|
|
|
|
|
|
||||
 |
|
 |
||
|
|
||||
|
Click on images for larger format |
||||
Name derivation: |
||||
|
‘Cloaked flagellate’ – Chlamy- cloak or mantle; -monas flagellate (Gr.)
|
||||
Classification: |
||||
|
Chlamydomonas Ehrenberg 1833; 414 of 1,163 species descriptions are currently accepted taxonomically. However the genus appears to be polyphyletic so transfer of some species to other genera will have to be done in the future (Pröschold et al. 2001). Order Chlamydomonadales; Family Chlamydomonadaceae The genus is polyphyletic, as are several other genera in the order, as it is distributed in five or more lineages (Buchheim et al. 1990, Buchheim et al. 1996, Pröschold et al. 2001, Nakada et al. 2008).
|
||||
Morphology: |
||||
|
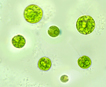
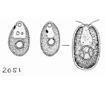
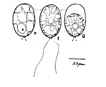
Chlamydomonas is single celled, and has an almost spherical cell wall around the
cytoplasm and centralized nucleus. They have two flagella extending out of
one side that propel them about. They contain an eyespot leading into ion
channels that react to changes in light allowing the cell to always move
closer to light (positive phototaxis). Near the flagella are two vacuoles
that are used for moving water into and out of the cell. Pyrenoids
are present in all Chlamydomonas
species.
|
||||
Similar genera: |
||||
|
Chloromonas spp. lack pyrenoids, but in all other morphological respects they resemble Chlamydomonas. There seems to be a controversy about which is the major widespread red snow algal genus, probably for lack of microscopic examination. (Guiry 2012). Synonym Protococcus C.Agardh 1824, including P. nivalis (a red snow alga), is not separated crom Chlamydomonas in Phycokey. Synonym Sphaerella S.C.Sommerfelt 1824
|
||||
Model genus in studies of PS, R, and PR: |
||||
|
Chlamydomonas is utilized in experimental studies of photosynthesis, respiration and photorespiration. An example is a review of electron transport studies in unicellular microalgae (Cardol et al. 2011) involving interactions between chloroplasts, mitochondria and peroxisome, and the balancing act of ATP energy and NADPH reducing power. Linear and cyclic electron flow (LEF, CEF) as well as the effect on pH changes are also discussed. Another excellent review explores current understanding of electron transport during anaerobiosis (Hemschemeier and Happe 2011) and flexibility in e-transport (McDonald et al. 2011). Further, the carbon concentrating mechanism (CCM) is an adaptation of photosynthetic organisms to photosynthesis rate-limiting levels of Ci (inorganic carbon as CO2, HCO3-1 and CO3-2). Major reasons for Ci limitation is the low supply in the atmosphere, and the slow rate of diffusion underwater (10-4 the atmospheric diffusion rate). Another factor is the relatively low affinity of Rubisco for Ci. Transport of ions (e.g. HCO3-1) across membranes (e.g. outer cell membrane, chloroplast membrane) requires a protein carrier. Accumulated cellular HCO3-1 is dehydrated to CO2 by carbonic anhydrase. Because CCM is an energy consumer, it is usually turned off in high concentrations of Ci. Spalding (2008) published an excellent review of CCM by active in Chlamydomonas . Reactive oxygen species (ROS) in Chlamydomonas such as H2O2 (peroxide) are recognized as signaling molecules (Shao et al. 2008). |
||||
Use: |
||||
|
Chlamydomonas reinhardtii is a potential species for production of biofuel as it is able to produce oil instead of starch when genetically modified (Spalding 2011). Apparently culture biomass is increased ~ 50% over wild type cells in the presence of high CO2 concentrations by keeping the CO2 uptake genes (LCIA and LCIB) active (normally they switch off under those conditions). Chemical energy storage in chlorophyceae is mainly starch. Green mutants unable to produce starch shunt their photosynthate toward triglycerides, demonstrating a competition between the two storage products (Li et al. 2010.. Most non-green microalgae store lipids rather than starch.
|
||||
Habitat: |
||||
|
Chlamydomonas can survive in many habitats. They can be found in marine, freshwater, ice, and sometimes soil situations. Due to their eyespot and ion channels that register light changes they are most often found as close to the light as they are able. Some species of Chlamydomonas have even been found in thriving in extremely cold or extremely acidic conditions.
|
||||
References: |
||||
|
Buchheim, M.A., M. Turmel, E.A. Zimmer and R.L. Chapman 1990. Phylogeny of Chlamydomonas (Chlorophyta) based on cladistic analysis of nuclear 18S rRNA sequence data. Journal of Phycology 26:689-699. Buchheim, M.A., C. Lemieux, C. Otis, R.R. Gutell, R.L. Chapan and M. Turmel. 1996. Phylogeny of the Chlamydomonadales (Chlorophyceae): A comparison of ribosomal RNA gene sequences from the nucleus and the chloroplast. Molecular Phylogenetic Evolution 5:391-402. Cardol, P., G. Forti and G. Finazzi 2011. Regulation of electron transport in microalgae. Biochimica et Biophysica Acta 1807:912-918. Guiry, M.D. and G.M. Guiry 2012. AlgaeBase. World-wide electronic publication, National University of Ireland, Galway. http://www.algaebase.org; searched on 22 December 2012. Hemschemeier, A. and T. Happe 2011. Alternative photosynthetic electron transport pathways during anaerobiosis in the green alga Chlamydomonas reinhardtii. Biochimica et Biophysica Acta 1807:919-926. McDonald, A.E., A.G. Ivanov, R. Bode, D.P. Maxwell, S.R. Rodermel and N.P.A. Hüner 2011. Flexibility in photosynthetic electron transport: The physiological role of plastoquinol terminal oxidase (PTOX). Biochimica et Biophysica Acta 1807:954-967. Nakada, T., K. Misawa and H. Nozaki 2008. Molecular systematic of Volvocales (Chlorophyceae, Chlorophyta) based on exhaustive 18S rRNA phylogenetic analyses. Molecular Phylogenetic Evolution 48:281-291. Pröschold, T., Marin, B., Schlösser, U.W. & Melkonian, M. (2001). Molecular phylogeny and taxonomic revision of Chlamydomonas (Chlorophyta). I. Emendation of Chlamydomonas Ehrenberg andChloromonas Gobi, and description of Oogamochlamys gen. nov. and Lobochlamys gen. nov. Protist152: 265-300, 7 figs, 5 tables. Shao, N., C.F. Beck, S.D. Lemaire and A. Krieger-Liszkay 2008. Photosynthetic electron flow affects H2O2 signaling by inactivation of catalase in Chlamydomonas reinhardtii. Planta 228:1055-1066. Spalding, M.H. 2008. Microalgal caron-dioxide=condentrating mechanisms: Chlamydomonas inorganic carbon transporters. J. Exp. Botany 59(7):1463-1473. Additional online resources: http://www.biology-resources.com/chlamydomonas-01.html Sept 15, 2010 http://www.jstor.org/stable/2482218 Sept 15, 2010 |
||||
|
Home / Chlorophyceae / Unicells / Flagellated / Chlamydomonas |
||||