|
Home / Diatoms / Pennate / Unicells / Biraphes / Asymmetric / Amphora |
||||
|
|
||||
|
|
||||
|
|
||||
 |
 |
|
|
 |
|
|
||||
|
|
|
|||
|
|
||||
 |
||||
|
Click on images for larger format |
||||
Name derivation: |
||||
|
Greek amphor meaning a two eared pitcher or flask. |
||||
Classification: |
||||
Amphora Ehrenberg ex Kutzing 1844; 306 of 1161 species descriptions are currently accepted taxonomically (Guiry and Guiry 2014).Order Thalassiophysales; Family CatenulaceaeEarly classification by Gregory (1857) divided Amphora into two groups of species on the basis of whether the frustule was ‘simple' (i.e. the girdle region appears more or less structureless and without striae in LM) or ‘complex' (the girdle bears transverse striations parallel to the pervalvar axis, as in Amphora commutata, or well-marked longitudinal striations). In contrast, Smith (1873) subdivided the genus according to the course of the raphe system (straight or incurved), the shape of the frustule (e.g. constricted or not; rostrate or not), the presence or absence of a stauros, and aspects of the valve striation (present or absent, as seen in LM, accompanied or not by longitudinal lines). Cleve (1895) combined aspects of both systems to produce nine subgenera (Sato et al., 2013) Taxon distributions were at least partly explained by environmental variables. Amphora had higher abundance at high pH and nutrient levels, while other diatoms were found at higher abundance at low pH and nutrient levels. However, relatively little is known about why certain diatom taxa are present in certain habitats (Kahlert and Gottschalk, 2014) |
||||
Morphology: |
||||
|
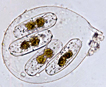
Solitary cells that can be motile and almost always occur in girdle view. The cells appear elliptical with flat truncate ends. The cell appears as a large slice of orange. The valves are asymmetrical and are sometimes smaller or constricted at each end of the cell. Both raphes lie on the same side of the valve. There are generally two or more plastids that occur in different positions throughout the cell. |
||||
Similar genera: |
||||
|
|
||||
Photosynthetic Activity: |
||||
|
In natural ecosystems, the benthic microalgae at low tide can be affected by high irradiances and elevated temperatures concurrently. High irradiance usually compounds the effects of extreme temperatures, and a combination of both factors results in lower photosynthetic rates. Amphora coffeaeformis had maximum values of photosynthetic parameters at temperatures between 10°C and 15°C at high irradiance levels (1,100 and 500 μmol photons · m−2 · s−1), while samples incubated in darkness had the highest photosynthetic rates at 20°C, which is similar to their growth temperature (Salleh and McMinn, 2011). |
||||
Habitat: |
||||
|
Epiphytic on plants, stones and mud. A large mainly marine genus with relatively few species in freshwater.
|
||||
Toxicity: |
||||
|
A strain from Canada was found to produce domoic acid. Other strains examined so far non-toxic. The precise identification of the Canadian strain has been questioned (Sala et al. 1998.) |
||||
|
|
||||
References: |
||||
|
Guiry, M.D. and G.M. Guiry 2014. AlgaeBase. World-wide electronic publication, National University of Ireland, Galway. http://www.algaebase.org; searched on 08 May 2014. Kahlert, M. and S. Gottschalk 2014. Differences in benthic diatom assemblages between streams and lakes in Sweden and implications for ecological assessment. Freshwater Science. Vol. 33(2): 655-669. University of Chicago Press. http://www.jstor.org/stable/10.1086/675727. Round, F. E., Crawford, R. M., Mann, D. G. 1990. The Diatoms. Cambridge University Press. New York, NY. Sala, S.E., E.A. Sar, and M.E. Ferrario. 1998. Review of materials reported as containing Amphora coffeaeformis (Agardh) Kützing in Argentina. Diatom Res. 13: 323-336. Salleh, S. and McMinn, A. 2011. The effects of temperature on the photosynthetic parameters and recovery of two temperate benthic microalgae, Amphora cf. coffeaeformis and Cocconeis cf. sublittoralis (Bacillariophyceae). Journal of Phycology 47(6):1413-1424. Sato, S., Nagumo Tamotsu, N and D. G. Mann (2013) Morphology and life history of Amphora commutata (Bacillariophyta) I: the vegetative cell and phylogenetic position. Phycologia: May 2013, Vol. 52, No. 3, pp. 225-238. |
||||


