|
Home / Cyanophyceae / Filaments / Unbranched / Tapered / Gloeotrichia or return to: |
||||
|
|
||||
|
|
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||
|
Click on images for larger format |
||||
Name derivation: |
||||
|
Filaments with viscous mucilage matrix.
|
||||
Classification: |
||||
Gloeotrichia J.Agardh ex Bornet and Flahault 1886; 17 of 29 species descriptions are currently accepted taxonomically (Guiry and Guiry 2013).Order Nostocales; Family NostocaceaeIn PhycoKey you can find Gloeotrichia either as a filament or a colony because it is a colony of filaments.
|
||||
Morphology: |
||||
|
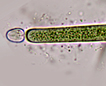
Spherical colonies of radiating straight trichomes (filaments without sheaths). Each trichome has an akinete as the basal cell near the center of the colony. Akinetes if present are adjacent the heterocyst. The primary morphology is trichomous (filamentous without sheaths), the secondary is colonial.
|
||||
Similar genera: |
||||
|
Rivularia is attached to a substrate and forms a morphologically similar colony of radiating, tapered filaments with basal heterocysts. |
||||
Habitat: |
||||
|
Freshwater plankton. In North America Gloeotrichia appears unexpectedly in many remote oligotrophic lakes during late summer and fall. During canoe trips through the Boundary Waters of Ontario, Manitoba and Minnesota I traversed several remote and pristine lakes in the undisturbed boreal forest watershed that had an abundance of Gloeotrichia colonies in the epilimnion. Recently Gloeotrichia was also found in 26 of 27 ‘low nutrient’ lakes in New England USA (Carey et al. 2012). Likely the colonies develop in the bottom waters where sediment mineralization releases a portion of its phosphate, then adjust their buoyancy with displacement of bacterioplasm by elongating gas vesicles and rise to the surface where they can be distributed horizontally by wind-driven water currents. Blooms form in mid to late summer due to this ‘recruitment’ from the sediments, as the benthic colonies rise relatively in synchrony, measured in inverted funnel traps at up to 104 colonies m-2 day-1 in Lake Sunapee, NH USA (Carey et al. 2014). Evidence that Gloeotrichia is meroplanktonic, spending part or most of the year in sediments, comes from mesocosm growth experiments at Lake Erken. While open-water (pelagic) colonies were increasing during July of 2000 – 2001, colonies in mesocosms (41 L and 300 L volume) were decreasing, even with additions of various combinations of nutrients (exception: addition of N, P and Fe) (Karlsson-Elfgren et al. 2005). The conclusion is that P-rich sediments enable colony growth and that increasing colony buoyancy during July brings them into the pelagic zone.
|
||||
|
Known Toxins |
||||
|
Microcystin LR was found in Gloeotrichia echinulata from Lake Sunapee, NH USA (Carey et al. 2007).
|
||||
References: |
||||
|
Bornet, É. and C. Flahault 1886. Revision des Nostocacées hétérocystées contenues dans les principaux herbiers de France. Annales des Sciences Naturelles, Botanique, Septième série 3: 323-381. Carey, C.C., J.F. Haney and K.L. Cottingham 2007. First report of Microcystin-LR in the cyanobacterium Gloeotrichia echinulata. Environmental Toxicology 22(3):337-339. Carey, C.C., H.A. Ewing, K.L. Cottingham, K.C. Weathers, R.Q. Thomas and J.F. Haney 2012. Occurrence and toxicity of the cyanobacterium Gloeotrichia echinulata in low-nutrient lakes in the northeastern United States. Aquatic Ecology 46:395-409. Carey, C.C., K.C. Weathers, H.A. Ewing, M.L. Greer and K.L. Cottingham 2014. Spatial and temporal variability in recruitment of the cyanobacterium Gloeotrichia echinulata in an oligotrophic lake. Freshwater Science 33(2):577-592. Graham, L. and L. Wilcox 2000. Algae. Prentice-Hall Karlsson-Eolfgren, I., P. Hyenstrand and E. Riydin 2005. Pelagic growth and colony division of Gloeotrichia echinulata in Lake Erken. Journal of Plankton Research 27(2):145-151. |
||||
|
Home / Cyanophyceae / Filaments / Unbranched / Tapered / Gloeotrichia or return to: |
||||